Durante siglos la investigación sobre la conciencia estuvo en manos de los filósofos, quienes no siempre se caracterizan por la búsqueda de evidencia empírica. Esto resultó en una sobreoferta de intuiciones junto con una escasez de medios para verificarlas. El estudio científico de la conciencia es como una historia de amor prohibido, una secuencia de desencuentros que postergaron hasta finales del siglo XX cualquier intento serio de abordar la conciencia a partir del estudio científico del cerebro. Los primeros psicólogos fundaron su disciplina alrededor del estudio de la conciencia, pero no disponían de suficientes herramientas metodológicas para establecer un cuerpo robusto y duradero de conocimiento sobre el tema. Lamentablemente, a medida que estas herramientas comenzaron a aparecer, los psicólogos redefinieron su objeto de estudio para abarcar únicamente el comportamiento observable de los organismos, lo cual naturalmente excluye la investigación sobre la conciencia.
El puntapié inicial para el estudio científico de la conciencia lo dio un físico (como yo) que, desencantado con los problemas por resolver en su propia disciplina (como yo), buscó un desafío en el entendimiento biológico de la conciencia (como yo) luego de haber obtenido el Premio Nobel de Fisiología de 1962 por determinar la estructura molecular del ADN (se terminaron las coincidencias). Francis Crick no fue el único científico que decidió probar suerte con el problema de la conciencia luego de ganar su Premio Nobel. Gerald Edelman (conocido por su trabajo sobre el sistema inmune) también se pasó al bando de la conciencia luego de su galardón en 1972, y Sir John Eccles formuló algunas escandalosas teorías dualistas sobre mente y cerebro luego de su paso por Estocolmo en 1963.
El hecho de que todos estos científicos hayan alcanzado el pináculo del reconocimiento antes de dedicarse a entender la conciencia es una muestra de la resistencia que existió durante la mayor parte del siglo XX a siquiera admitir que se trataba de un objeto de estudio legítimo. Antes del trabajo publicado en 1990 por Francis Crick y Christof Koch, titulado “Hacia una teoría neurobiológica de la conciencia”, difícilmente un científico hubiese decidido dedicarse a este problema, principalmente porque los prejuicios de la comunidad científica habrían resultado en enormes dificultades para conseguir financiamiento, montar un laboratorio, viajar y exponer sus resultados en congresos internacionales. Como en Argentina todas esas dificultades ya están dadas automáticamente por los crónicos problemas presupuestarios, independientemente del tema de estudio, decidí que no me hacía falta esperar a tener un Premio Nobel para investigar sobre la conciencia.
Para entender la enorme influencia del artículo publicado por Crick y Koch primero es necesario recorrer la historia del siglo XX para responder la siguiente pregunta: ¿por qué estaba mal visto estudiar la conciencia y qué fue lo que cambió en ese momento?
Tal como anticipamos al comienzo del capítulo, la conciencia fue investigada intermitentemente desde finales del siglo XIX. Dado que todos tenemos una conciencia y la podemos examinar aplicando nuestras facultades introspectivas, la posibilidad de abordar el problema parece estar al alcance de todo el mundo. Pero la introspección es un instrumento engañoso, al menos si no se la complementa con mediciones objetivas sobre lo que sucede en el cerebro. A principios del siglo XX, los primeros psicólogos (William James, Wilhelm Wundt, Edward Titchener, y otros) se embarcaron en el programa de investigar sistemáticamente la estructura de su mente mediante la introspección, aunque no siempre consiguieron ponerse de acuerdo. Enfrentados a conceptos tales como “los átomos del pensamiento que constituyen la experiencia”, distintos introspeccionistas encontraron respuestas disímiles; por ejemplo, algunos concluyeron que los elementos fundamentales de la experiencia consciente se contaban en las decenas, mientras que otros llegaron a contar cientos o miles. El problema, por supuesto, es que es muy difícil acordar el significado de conceptos abstractos como “átomos del pensamiento” (y, muy posiblemente, tampoco tenga sentido hacerlo). Otro problema es que, por la propia naturaleza de su método, los introspeccionistas únicamente eran capaces de acceder a la experiencia consciente. Pero el conocimiento científico de la conciencia no depende tan solo de estudiar los comportamientos que requieren conciencia, sino también aquellos que no la requieren.Así es como descubrimos, por ejemplo, que la conciencia se asocia al procesamiento secuencial de la información en el cerebro, mientras que actividades que pueden ejecutarse automáticamente y por fuera de la conciencia (por ejemplo, tocar el piano, andar en bicicleta, identificar caras en una escena visual) se sirven de circuitos neuronales que procesan la información en paralelo.
El introspeccionismo fue finalmente descartado por culpa de sus inconsistencias y limitaciones, aunque es importante reconocer que también fue el marco de experimentos completamente revolucionarios para su época. Wilhelm Wundt, por ejemplo, había desarrollado un sofisticado pensamiento psicológico sobre la relación entre mente y materia, en gran medida inspirado por Gottfried Leibniz. En su laboratorio de psicología experimental en Leipzig, Alemania (el primer laboratorio de psicología experimental de la historia), Wundt realizaba experimentos controlados en los que medía tiempos, presentaba estímulos, y registraba reacciones. Un rasgo único de los experimentos de Wundt (y otros introspeccionistas) era considerar la impresión subjetiva de los sujetos como un dato científico. Por ejemplo, Wundt intentaba medir la velocidad del pensamiento enfrentando a una persona con un reloj de péndulo y pidiéndole que reportara la posición del mismo durante distintas etapas de sus procesos mentales. Los introspeccionistas consideraban que la expresión de los sujetos sobre sus propios estados mentales constituía un dato valioso y que merecía ser explicado, aunque podían llegar a conclusiones contradictorias cuando únicamente se apoyaban en la introspección y desestimaban la necesidad de complementarla con mediciones objetivas.
El introspeccionismo fue sepultado completamente por una escuela filosófica y científica llamada comportamentalismo (o conductismo), según la cual el objetivo de la psicología es relacionar los estímulos que recibe un organismo con su comportamiento. No hay lugar en el comportamentalismo para la introspección, el estudio de las experiencias subjetivas o incluso una vida mental interna. Esta escuela de pensamiento estaba influenciada fuertemente por el positivismo y su requerimiento de operacionalizar los objetos de estudio científicos, es decir, de determinar operaciones que permitan medirlos y cuantificarlos. Las inconsistencias del introspeccionismo fueron interpretadas como la imposibilidad de operacionalizar la experiencia subjetiva. En cambio, el estudio del comportamiento resulta de forma natural en una disciplina científica más pragmática, reproducible y cuantificable, que dominó por completo la psicología experimental durante la mayor parte del siglo XX.
El estudio de la conciencia es posiblemente lo más absurdo que puede existir para un comportamentalista convencido. La conciencia es un objeto de estudio nebuloso e indeterminado. Cuantificar fielmente su estructura mediante la introspección (tal como intentaron los introspeccionistas) parece ser equivalente al fracaso. Quizás el mayor obstáculo de todos es que la conciencia parece desarrollarse en una dimensión independiente de la del comportamiento. Yo puedo cerrar mis ojos y conjurar las más vívidas imágenes mentales sin dar señales externas sobre los contenidos de mi conciencia. ¿Cómo sería posible, entonces, medir objetivamente un comportamiento unívocamente asociado a determinadas experiencias conscientes?
Así como el introspeccionismo fue sepultado por el enorme éxito del comportamentalismo, este fue condenado en gran medida a desaparecer por el enfoque de la psicología experimental que sigue siendo dominante hasta el día de hoy: el cognitivismo. Conceptualizar un organismo como una caja negra que transforma estímulos en comportamientos mediante ciertas reglas a determinar es una primera aproximación útil, pero también es profundamente limitada. Podemos ilustrar estas limitaciones con una anécdota sobre la vida de John von Neumann, el brillante matemático húngaro, arquitecto de la bomba atómica y de la primera computadora digital.
Von Neumann era famoso por la velocidad de su pensamiento y por su memoria casi perfecta. En una época fecunda en inmigrantes húngaros con habilidades cognitivas excepcionales (conocidos como “los marcianos” –sin relación alguna con los científicos extraterrestres que aparecieron en el primer capítulo–), el intelecto de Von Neumann sobresalía entre todos ellos y era una continua fuente de asombro para sus amigos y colegas.
Según cuenta la anécdota, el físico Max Born le propuso en una ocasión el siguiente acertijo a Von Neumann:
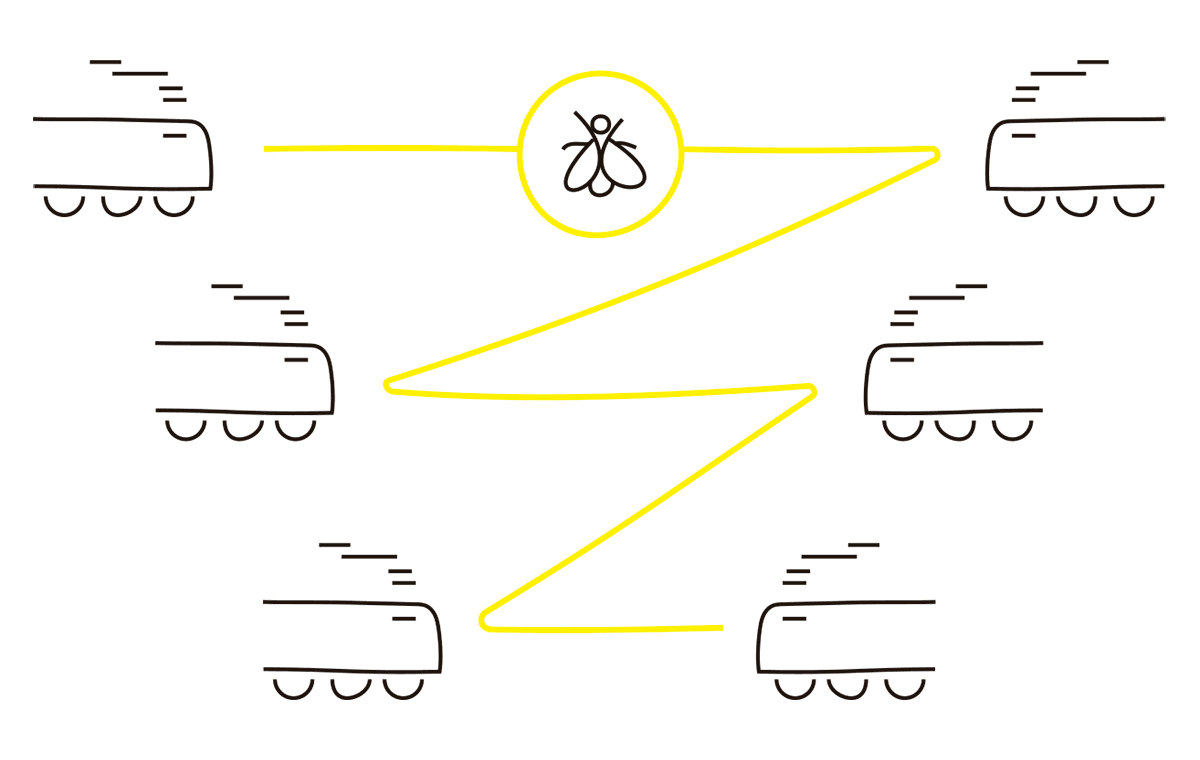
Dos trenes separados por 40 km se acercan en rumbo de colisión a una velocidad de 20 km por hora. Mientras se acercan, una pequeña mosca vuela a una velocidad de 60 km por hora desde la locomotora de uno de los trenes hacia la del otro, y luego regresa, repitiendo el vuelo hasta que los trenes chocan y la mosca es aplastada. La pregunta es: ¿qué distancia recorrió la mosca hasta que fue aplastada entre los trenes?
Este acertijo es divertido porque existen dos maneras de resolverlo, una fácil y otra muchísimo más complicada. La solución complicada consiste en calcular la distancia que recorre la mosca en la primera parte del trayecto, desde una locomotora a la otra (teniendo en cuenta que la locomotora opuesta se está aproximando constantemente). Una vez obtenido el resultado, lo guardamos en la memoria. Luego, calculamos la distancia que recorre la mosca al regresar a la otra locomotora, y también guardamos este número en la memoria, y así sucesivamente. El resultado final se obtiene sumando todas las distancias que calculamos. El problema es que no importa qué tan cerca estén las locomotoras, tenemos que sumar la distancia que recorre la mosca entre ellas, lo que significa que tenemos que calcular y sumar una cantidad infinita de términos. En la jerga matemática, diríamos que tenemos que sumar una serie infinita. Una pesadilla matemática, incluso teniendo papel, lápiz y tiempo a disposición:
FIG. 3.1 EL ACERTIJO DEL TREN Y LA MOSCA
Por otro lado, la solución fácil es realmente muy fácil. El truco es darse cuenta de que los trenes separados por 40 km se aproximan a 20 km por hora el uno hacia el otro, por lo que exactamente en una hora se encontrarán en el punto medio de los 40 km iniciales que los separaban. Como la mosca vuela siempre a 60 km por hora, y vuela por una hora hasta que los trenes se chocan, la respuesta es muy sencilla: la mosca recorre 60 km antes de ser aplastada por el choque.
Según la anécdota, luego de que Max Born le planteara el acertijo a Von Neumann, obtuvo de forma casi instantánea la respuesta correcta: “La mosca recorre 60 km”. Desencantado, Born le reprochó: “No tiene gracia, conocías el truco”, a lo que Von Neumann respondió, confundido: “¿Truco? ¿Qué truco? Lo único que hice fue sumar la serie infinita”.
El punto es el siguiente: tratar a Von Neumann como una caja negra que transforma estímulos en respuestas evade por completo la cuestión esencial de cómo se realiza esa transformación. Claramente existen muchas formas distintas de llegar a la solución del mismo problema, y esas formas pueden ser dramáticamente diferentes. En particular, el tiempo no es el único factor que tenemos que tener en cuenta (como ilustra la equivocación de Max Born respecto de los procesos mentales de Von Neumann). El problema del comportamentalismo es que rechaza el estudio de los estados internos de la caja negra. Únicamente mide variables externas al sistema y, por lo tanto, es incapaz de teorizar sobre los distintos procesos computacionales posibles que ocurren dentro del cerebro para vincular un estímulo con un determinado comportamiento.
La respuesta del cognitivismo al comportamentalismo está, en gran medida, inspirada en la computadora digital (justamente, desarrollada por John von Neumann). Por un lado, una computadora digital parece ser solamente una caja negra que transforma ciertos inputs (lo que tipeamos o indicamos con clics) en outputs (lo que aparece en el monitor). Pero todos sabemos que dentro de esa caja negra existen instrucciones (que implementan algoritmos) que se encargan de realizar esa transformación. Incluso hay algoritmos más rápidos y eficientes que otros para resolver el mismo problema (similar a las dos formas de resolver el acertijo de la mosca). Por lo tanto, es esencial describir el estado interno de una computadora para entender por completo su funcionamiento. El legado de científicos como Alan Turing o John von Neumann es un marco teórico para describir esos estados internos, lo que coloquialmente llamamos un programa. Un programa es una serie de instrucciones que ejecutan distintas funciones e interactúan con componentes físicos de la computadora, tales como la memoria y la unidad de procesamiento central (CPU), entre otros. La implementación de un programa resulta en una cadena de eventos causales que se dispara cuando nosotros damos un input determinado: a partir de entonces, la computadora implementa secuencialmente ciertas funciones, que, a su vez, implementan otras funciones, y así sucesivamente. Finalmente, obtenemos nuestra respuesta en la pantalla del monitor.
El cognitivismo propone precisamente una analogía de la mente como instrucciones en una computadora: un conjunto de funciones caracterizadas por el papel que juegan dentro de una red de causas y efectos. En otras palabras: el cognitivismo reemplaza la caja negra de los comportamentalistas por un programa que vincula el input con el output, y es mediante la naturaleza de ese programa que alcanzamos un conocimiento más profundo sobre la mente y su relación con el cerebro. Algunas de las funciones de este programa nos son familiares: lenguaje, memoria, atención, toma de decisiones y muchas otras. Nuestra cognición es el conjunto de esas funciones y los modos en que se entrelazan causalmente para dar origen a nuestro comportamiento. Esta analogía computacional de la mente humana está inspirada, claramente, en la posición filosófica del funcionalismo.
Los ecos del comportamentalismo todavía resonaban por las aulas, pasillos y centros de fotocopiado de las universidades cuando Crick y Koch invirtieron la tortilla epistemológica con la publicación de su trabajo. A partir de ese momento, el estudio neurocientífico de la conciencia comenzó a florecer aceleradamente, de manera casi exponencial. En apenas trece páginas, Crick y Koch expusieron una única idea que dio origen al estudio moderno de la conciencia; una idea que sigue influenciando, explícita o implícitamente, el pensamiento de todos los neurocientíficos interesados en el problema de la conciencia. Antes de explicar la idea seminal de Crick y Koch, es muy instructivo leer algunos pasajes textuales de su artículo.
El tono ligeramente mesiánico del trabajo nos sugiere la inmensa barrera cultural y social que estaban intentando atravesar. Lo mismo sucede con la idea central que proponen, de tal simplicidad que, estando durante décadas a la vista de todos, su sistemática omisión puede explicarse únicamente por el Zeitgeist dominante en la ciencia del siglo XX:
Es notable que la mayor parte del trabajo en las ciencias cognitivas y las neurociencias no menciona la conciencia en absoluto, especialmente dado que uno consideraría la conciencia como el mayor problema que confronta la idea de que la mente posee una base neuronal, y que en el momento presente parece profundamente misteriosa para la mayoría de las personas. Esta actitud es en parte un legado del comportamentalismo, y en parte se debe a que la mayoría de quienes trabajan en esta área no logran concebir una forma útil de aproximarse al problema. En los últimos años, han aparecido varios libros que consideran la pregunta de forma directa, pero la mayoría de estos libros fueron escritos desde una perspectiva funcionalista y, por lo tanto, tienen poco para decir sobre las neuronas y la maquinaria del cerebro.
En este elegante párrafo inicial, Crick y Koch afirman que:
La conciencia es el mayor problema de la neurociencia y, sin embargo, nadie parece estar estudiándolo.
El comportamentalismo, con su tendencia a reemplazarnos por cajas negras sin vida interior, tiene algo de culpa de esta situación.
Incluso logrando escapar de la perspectiva del comportamentalismo, la realidad es que a nadie se le ocurre cómo avanzar en este problema.
Algunas personas creen que se les ha ocurrido, pero están equivocadas. Esto es porque sus explicaciones se basan únicamente en las funciones del cerebro; en otras palabras, en considerar que la mente es comparable con una secuencia de instrucciones de un programa de computadora. Pero ¿dónde está la conciencia dentro de esas instrucciones? Para Crick y Koch no es obvio que la conciencia pueda conceptualizarse de esta manera, es decir, en términos funcionales.
Según ellos, el problema es haber considerado la conciencia como un problema para la psicología experimental, la clase de problema que respondemos mediante explicaciones sobre la mente y sus funciones. Por el contrario, para Crick y Koch la conciencia es un problema para la neurociencia: la explicación que debemos encontrar es en términos de neuronas disparando y neurotransmisores siendo liberados en vesículas sinápticas, sin referencia a las funciones mentales que identifican los psicólogos y científicos cognitivos.
Podemos contrastar ambas posiciones inventando dos explicaciones alternativas para la conciencia:
“La conciencia es aquella función intermediaria que vincula nuestra percepción y nuestra memoria con nuestra capacidad para tomar decisiones, actuar y expresarnos por medio del lenguaje”.
“La conciencia es el resultado de neuronas en el tálamo y en la capa más externa de la corteza que sincronizan sus disparos de forma colectiva mediante la acción de neurotransmisores excitatorios, tales como el glutamato”.
Para Crick y Koch, una explicación del primer tipo nunca podría ser realmente informativa sobre la conciencia: inventar una función llamada conciencia no es suficiente para entender cómo puede ocurrir tal cosa en el cerebro.
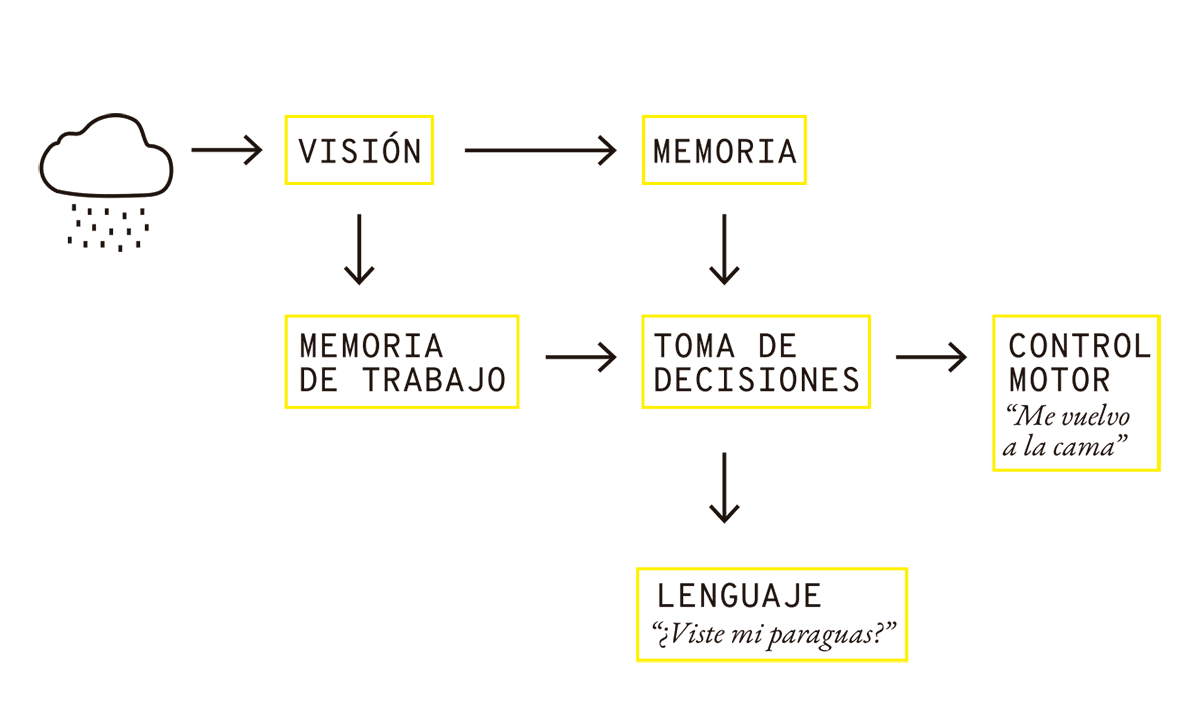
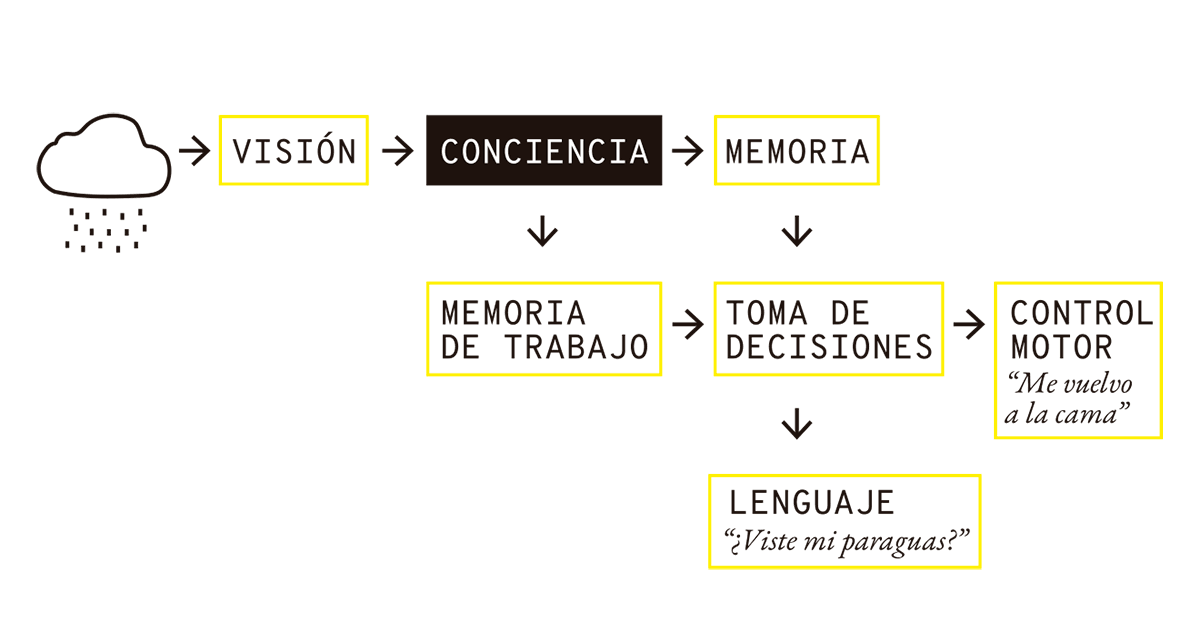
Pensemos en otro ejemplo: alguien se levanta durante una mañana lluviosa y mira por la ventana. Algunos segundos después, decide o bien volver a su cama, o bien preguntar dónde quedó su paraguas para poder salir a trabajar. ¿Cómo podemos explicar lo que sucedió?
Primero, la persona utilizó su sistema visual para formar una imagen de las nubes lluviosas y reconocerlas como tales, y envió esa información a dos módulos distintos. El módulo “memoria de trabajo” sirve para mantener la imagen de las nubes durante un tiempo para ser considerada por otros procesos (es una suerte de memoria RAM). Al mismo tiempo, la información accede a una memoria más consolidada que contiene datos relevantes como qué hay que hacer hoy en el trabajo, cuántas veces falté en lo que va del mes, detalles de la relación con mi jefa, etc. En algún momento, esas consideraciones convergen en la toma de una decisión, y dependiendo de esa decisión, puede activarse el control motor que desplaza a la persona nuevamente a la cama, o una verbalización del estilo: “¿Viste mi paraguas?”.
FIG. 3.2 DE LA LLUVIA A LA DECISIÓN
Dejemos de lado el problema de si esto constituye una explicación convincente y vamos al punto central que preocupaba a Crick y Koch: en ningún momento encontramos un rectángulo que diga “conciencia” en el diagrama que explica el comportamiento del sujeto. Y si lo encontrásemos, ¿de qué nos serviría? El problema es que ese rectángulo no es como los otros porque no cumple un rol computacional claro. Podemos estudiar el control motor en el cerebro relacionando la actividad de neuronas con la contracción de ciertos músculos; la memoria, con la capacidad para utilizar información del pasado en el momento presente; y la visión, con el procesamiento de fotones que inciden en la retina y la extracción de datos relevantes a lo largo de la jerarquía visual. Pero ¿la conciencia? Poner una caja que diga “conciencia” en este diagrama es equivalente a posponer la determinación de su rol en el procesamiento de la información en el cerebro.
Enfrentados con esta dificultad, Crick y Koch vienen a salvar el día y proponernos una forma distinta de abordar el problema. Pero antes de hacerlo, dedican varios párrafos a considerar y descartar distintas teorías cognitivas de la conciencia (por ejemplo, la teoría de que la conciencia es una especie de “sistema operativo” del cerebro). La ironía es que la teoría más dominante y completa de la que disponemos sobre la conciencia en la actualidad es, justamente, una teoría formulada desde la perspectiva del cognitivismo. Pero, tal como vamos a ver en el próximo capítulo, incluso esa teoría le debe mucho a la idea fundamental y transformadora de Crick y Koch.
La idea fundamental del artículo es la siguiente: el objetivo es encontrar el mínimo conjunto de eventos físicos en el cerebro que se corresponden con una determinada experiencia consciente. A este conjunto lo denominan correlatos neuronales de dicha experiencia consciente. Por “eventos físicos” queremos decir cosas tales como disparos de neuronas o liberaciones de neurotransmisores, entre otros procesos medibles en el laboratorio. Crick y Koch plantean, a modo de ejemplo, las siguientes preguntas: ¿dónde están las neuronas cuya actividad corresponde al mínimo indispensable para que ocurra una experiencia consciente? ¿Qué tipo de neuronas son? ¿Cómo están conectadas entre sí y con las demás neuronas? ¿Hay algo especial en la forma en que disparan a lo largo del tiempo?
El problema no está planteado en términos funcionales; en principio, no interesa la función que cumplen dichos correlatos neuronales. Tampoco está planteado en términos causales, porque no sabemos si esa actividad neuronal causa la conciencia, únicamente sabemos que está correlacionada con esta. Además, no nos interesan las neuronas cuya actividad sea optativa respecto de la conciencia, sino el mínimo conjunto, el más indispensable, aquel en el cual remover una única neurona destruiría la capacidad de tener esa experiencia consciente.
El mensaje de Crick y Koch por fin queda claro: si la conciencia resulta demasiado misteriosa, encontremos, entonces, eventos físicos en el cerebro que se correspondan completamente con la conciencia. Cada vez que esos eventos físicos ocurran, la experiencia consciente se manifestará. Y si la experiencia consciente no se manifiesta, será porque esos eventos físicos no ocurrieron.Una lectora o lector atento notará que esta perspectiva se encuentra alineada con la teoría de la identidad psiconeural, según la cual la conciencia se identifica con los procesos físicos que suceden en el cerebro. En cambio, le da la espalda a la perspectiva funcionalista. Pensando en el problema de esta forma, no importa qué tan misteriosa sea la conciencia, porque sus eventos físicos asociados son solo eso: eventos físicos. Hacemos física desde hace siglos, quizás milenios, así que si logramos encontrar los correlatos neuronales de la conciencia, entonces por fin lograremos traducir el problema a un lenguaje que podemos entender, procesar y analizar.
Supongamos que uno de los científicos marcianos del primer capítulo visita la Tierra y se encuentra con una computadora. Los científicos marcianos tienen una sólida formación en psicología y neurociencia, pero no son particularmente buenos en electrónica. Así, el visitante marciano siente intriga y decide tratar de entender cómo funciona la computadora. La función de las computadoras es computar, por lo que el marciano busca comprender cómo es que el objeto que tiene delante es capaz de dar respuestas a problemas tales como calcular la raíz cuadrada de 7 o encontrar todas las soluciones de una ecuación polinomial.
Resulta que todos los marcianos leen el trabajo publicado por Crick y Koch cuando están en preescolar. Por lo tanto, este marciano en particular recuerda su tesis central y decide aplicarla para entender cómo funciona la computadora; es decir, intenta encontrar el conjunto mínimo de eventos físicos asociado con su funcionamiento. La impresora, por ejemplo, puede ser descartada. El teclado, también (podemos ingresar el input inyectando corriente en ciertas partes del circuito impreso), así como el monitor (podemos leer el resultado midiendo otras corrientes en otras partes del circuito). Dependiendo del cómputo en cuestión, remover el disco rígido podría ser demasiado (quizás la memoria RAM no alcance). Pero en algunos casos, es seguro que podremos remover el disco rígido y también grandes porciones de la memoria RAM. La CPU parece indispensable para cualquier cómputo, pero si sabemos muchísimo de microelectrónica, podemos descubrir que para ciertos cómputos no circula corriente por algunos circuitos de la CPU, y entonces también podemos removerlos.Podemos imaginar, por ejemplo, que hay una parte del circuito dedicada a “llevarse dígitos”. En ese caso, esa parte del circuito es optativa para la multiplicación de 12 por 13, pero no para 17 por 13. Así, el marciano logra descubrir el mínimo conjunto de piezas, cables y circuitos necesarios para que un cómputo ocurra. Ese conjunto no es siempre el mismo: puede cambiar dependiendo del cómputo. Pero seguro que para cada cómputo específico es posible encontrar un mínimo conjunto asociado.
Los seres humanos no necesitamos pasar por ese proceso para entender cómo computa una computadora, porque nosotros somos quienes las diseñamos y las construimos. La situación es muy distinta en el caso del cerebro. La frase “Lo que no puedo crear, no lo puedo entender” estaba escrita en el pizarrón del famoso físico Richard Feynman al momento de su muerte, y refleja el abismo conceptual que existe entre entender el funcionamiento del cerebro y la conciencia, y entender el funcionamiento de una computadora.
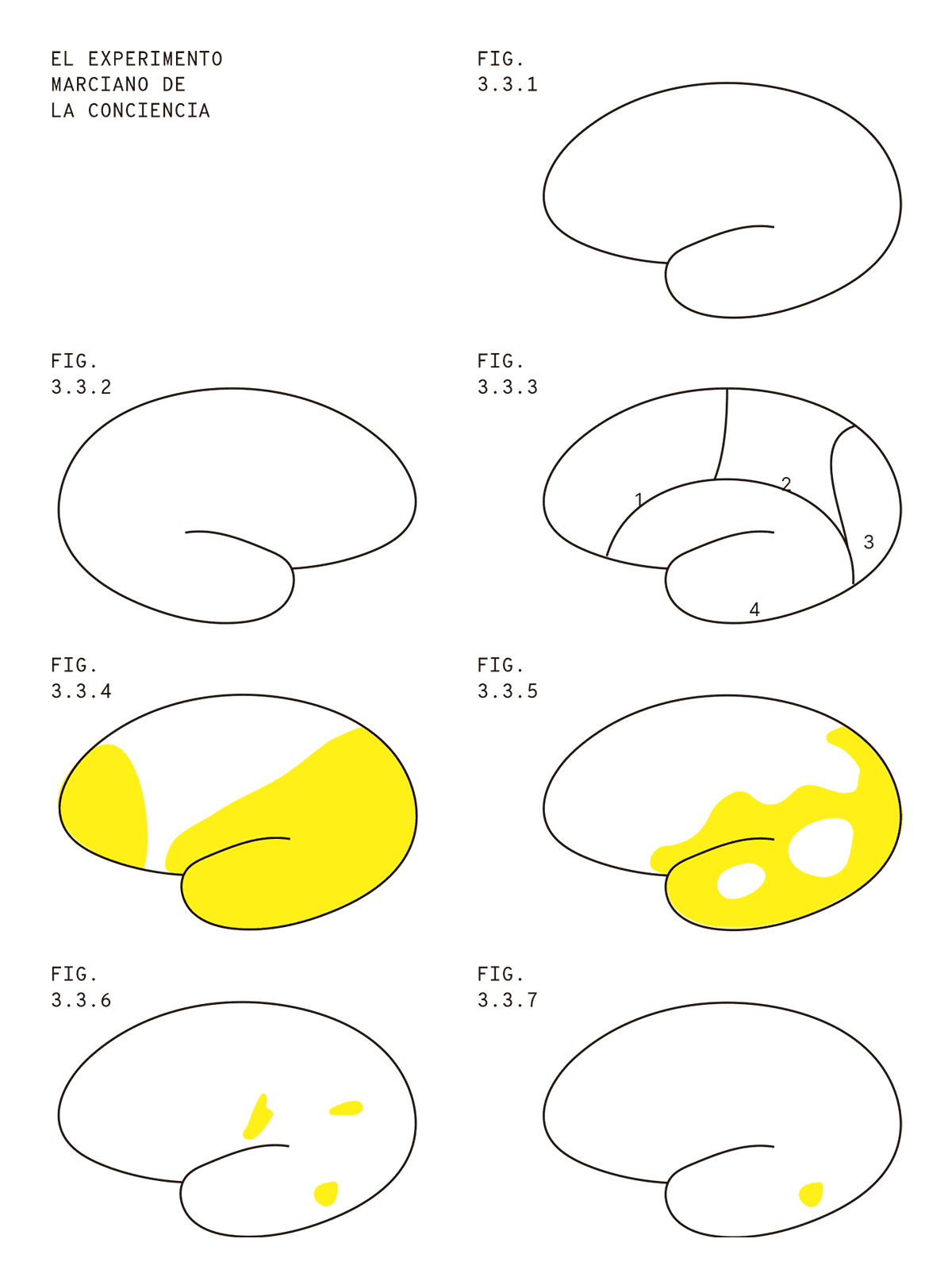
Ahora que ilustramos la idea de Crick y Koch para el caso de una computadora, busquemos entender cómo se traslada al cerebro y la conciencia. Así dibujo yo los cerebros (Fig. 3. 3.1). Este en particular (Fig. 3.3.2) es un cerebro visto de costado. Podemos verlo desde el costado opuesto y encontrar algo muy similar, sobre todo porque yo dibujo lo mismo, pero al revés.
Los cerebros también se pueden ver desde arriba y desde el frente, pero como no me sale dibujarlos desde esas perspectivas, vamos a limitarnos a verlos de costado (si necesitan dibujos más precisos de cerebros, en internet está lleno). Podemos dividir el cerebro en distintas zonas y asignarle un número a cada una (Fig. 3.3.3).
El cerebro tiene una superficie arrugada, con hendiduras llamadas surcos (que tampoco sé dibujar muy bien). Las áreas numeradas están separadas entre sí por grandes surcos. El área número 1 se llama lóbulo frontal, el área número 2 se llama lóbulo parietal, el área número 3 se llama lóbulo occipital y el área número 4 se llama lóbulo temporal. El lóbulo frontal está en la parte delantera del cerebro (un poco por encima de los ojos) y el lóbulo occipital está en la parte trasera del cerebro.
Después de su experiencia con la computadora, el científico marciano regresa a Marte con algunos “voluntarios”, decidido a hacer una demostración práctica de la idea de Crick y Koch para su clase “Historia de la ciencia terrícola”. El marciano ata al sujeto experimental a una silla y le presenta distintas imágenes, por ejemplo, la imagen de un perro labrador, y luego pide al sujeto indicar verbalmente qué está viendo (“perro labrador” sería, en este caso, la respuesta correcta). Mediante técnicas avanzadas de neuroimágenes, logra generar un mapa de todas las neuronas del cerebro que están respondiendo de una forma u otra durante el experimento (Fig. 3.3.4).
Ahora, el científico marciano utiliza un escalpelo láser de alta precisión para destruir distintas zonas del cerebro del sujeto experimental (Fig. 3.3.5). Imitando el procedimiento llevado a cabo con la computadora, demuestra que puede remover una buena parte del cerebro sin cambios mayores en relación con la experiencia consciente. Esto no significa que el comportamiento de la persona permanezca igual, sino que (de acuerdo a su comportamiento y sus reportes) sigue experimentando la imagen de un perro labrador.En el cerebro humano hay neuronas asociadas a la regulación del sistema autónomo (respiración, regulación cardíaca, etc.). Podemos asumir que el marciano se encarga de mantener el cerebro adecuadamente oxigenado e irrigado luego de eliminar esas neuronas. También podemos imaginar que destruye los contenidos de la memoria excepto los mínimos indispensables para que la persona sepa que tiene que decir “es un perro labrador” cuando es consciente del perro labrador, o cualquiera sea el mensaje acordado con el experimentador. La persona no tiene por qué tener sentido de su identidad personal, ni de su cuerpo, ni de su presente, pasado o futuro, pero sí tiene que ser capaz de tener la experiencia visual de un perro labrador y de reportarla.
Recordemos: el científico marciano tiene el propósito de encontrar el mínimo conjunto de neuronas cuya activación es necesaria para que el sujeto tenga la experiencia consciente de ver un perro labrador.
En un determinado momento, el marciano aplica el escalpelo y el sujeto finalmente declara ser incapaz de ver al perro labrador. En realidad, el sujeto afirma estar ciego, porque no registra ningún tipo de experiencia visual. Pero el científico marciano recuerda su experiencia con la computadora en la Tierra. Incluso removiendo el teclado, fue posible introducir el input mediante la activación de ciertas terminales en el microprocesador de la computadora. De la misma manera, ahora el científico marciano utiliza un instrumento para activar ciertas neuronas y descubre que el sujeto experimental nuevamente está convencido de ver un perro labrador, aunque es lo único que puede ver, y solamente puede verlo cuando el científico marciano activa esas neuronas. La conclusión es que el marciano eliminó en algún momento el “teclado”: un grupo de neuronas cuya actividad era necesaria para comunicar el input visual a aquellas que forman la experiencia consciente. Pero esto se solucionó simplemente estimulando esas neuronas de forma directa. Ahora solo quedan tres pequeños grupos de neuronas intactas en el cerebro del sujeto (Fig. 3.3.6).
El experimento marciano sobre la conciencia
El marciano entonces destruye uno de esos tres grupos, de forma tal que, incluso con la estimulación correcta para generar la experiencia de “perro labrador”, el sujeto experimental permanece callado. Pero todos en el aula entienden lo que está pasando: así como no necesitábamos un monitor para leer el output del cómputo realizado por la computadora (porque podíamos registrarlo directamente de la corriente eléctrica en ciertas partes de un microcircuito), tampoco necesitamos neuronas que se encarguen del lenguaje, y por lo tanto, de vocalizar la frase “es un perro labrador”. Pero eso no significa que el sujeto no esté siendo consciente de la imagen visual de un perro labrador cuando es adecuadamente estimulado. Únicamente es incapaz de decirlo. Por lo tanto, resta encontrar un grupo de neuronas donde se represente lo que el sujeto está intentando decir y, a partir de ahí, será posible registrar la respuesta correcta luego de la estimulación.
En ese momento, el profesor marciano se dirige al aula y afirma con seriedad: “Entonces, lo último que nos queda hacer para encontrar los correlatos neuronales de la experiencia visual de un perro labrador es eliminar las neuronas de las cuales podemos decodificar el reporte verbal ‘es un perro labrador’. Porque no es necesario afirmar que uno está viendo un perro labrador para tener una experiencia consciente de un perro labrador. Así que voy a remover esas neuronas, y finalmente vamos a descubrir los correlatos neuronales de la experiencia consciente” (Fig. 3.3.7).
Inmediatamente, el aula estalla en carcajadas. Todos entienden, por supuesto, el callejón sin salida en el cual se metieron al perseguir la idea de Crick y Koch hasta sus últimas consecuencias. El callejón sin salida es el siguiente: no hay forma alguna de saber si el sujeto sigue teniendo una experiencia de un perro labrador cuando se estimulan las neuronas que quedaron.Hay un paralelo con una conocida historia sobre ciencia y reduccionismo. Un biólogo descubre que una araña se mueve cuando hace sonar una campana. Entonces, anestesia una de sus patas y descubre que sigue moviendo las patas restantes al sonar la campana. Luego, repite este proceso hasta que finalmente anestesia la última y octava pata. Llegado ese punto, la araña se queda quieta cuando suena la campana, así que el biólogo concluye que las arañas escuchan con sus patas. Podría haber pasado que las últimas neuronas que el marciano destruyó en realidad eran parte de lo mínimo indispensable para que ocurra la experiencia consciente de un perro labrador. Ahora lo único que toda la clase está viendo es un grupúsculo de neuronas que disparan cuando el marciano las estimula, pero nadie en el aula cree realmente que esas neuronas estén experimentando algo.
Un funcionalista podrá encontrar otro problema en el procedimiento llevado a cabo por el marciano. Cuando este reemplaza una región cerebral por una estimulación externa adecuada, omite explicar cómo y dónde se computa la estimulación que debe aplicarse. Puede que este cómputo sea sencillo cuando se reemplazan áreas periféricas del cerebro, pero sin duda se vuelve más y más complejo a medida que se avanza hacia regiones involucradas en funciones cognitivas más sofisticadas. Quizás a la larga la única manera de computar la estimulación sea mediante un sistema artificial muy similar (o incluso idéntico) al pedazo de cerebro removido. Pero en ese caso, ¿por qué diríamos que ese pedazo es opcional para la conciencia? Sin duda está involucrado, y aunque se trate de un sistema artificial, esto no implica que no pueda ser equivalente a una región cerebral desde el punto de vista funcional. Entonces, ¿cómo podemos delimitar los correlatos neuronales de la conciencia en el cerebro, cuando no podemos decidir si los sistemas artificiales que aplicamos para imitar la función de ciertas zonas del cerebro forman parte o no de dichos correlatos?
El proceso de encontrar los correlatos neuronales de la conciencia es muy extraño. Por un lado, requiere eliminar todo aquello que no sea necesario para que ocurra una experiencia consciente. Si algo es necesario, por el contrario, no debemos eliminarlo. La intuición nos dice que el habla, por ejemplo, no es necesaria, porque somos conscientes de cosas todo el tiempo sin tener que hablar sobre ellas. Lo mismo sucede, aparentemente, para las demás funciones cognitivas. Por otro lado, ¿realmente creemos que puede existir la percepción visual consciente de un perro labrador sin un sujeto que la perciba ni pueda hablar de ella, manifestándose cuando estimulamos un grupo de neuronas en el cerebro?
El proyecto de encontrar los correlatos neuronales de la conciencia no es una fantasía filosófica sobre científicos marcianos que ironizan acerca del pobre entendimiento humano. Se trata de un proyecto real, con experimentos reales que pueden interpretarse a la luz del hipotético experimento marciano de la sección anterior. Uno de los más famosos involucra un fenómeno conocido como rivalidad binocular. Si presentamos una imagen distinta a cada ojo, ¿qué es lo que vemos? Para algunos, la respuesta intuitiva es “una superposición de ambas imágenes”. Por ejemplo, si las imágenes que presentamos al ojo izquierdo y derecho son las siguientes:
FIG. 3.4.1 RIVALIDAD BINOCULAR: IMAGEN DISTINTA PARA CADA OJO
Una posible intuición es que la percepción visual consciente será la superposición de ambas figuras:
FIG. 3.4.2 INTUICIÓN: SUPERPOSICIÓN DE IMÁGENES
Sin embargo, esta intuición no es acertada. No vemos este símbolo (que coincide con la letra griega mayúscula “φ”). Lo que vemos en realidad es una alternancia de ambas imágenes: primero una, luego la otra, así sucesivamente. La alternancia ocurre más allá de nuestra voluntad: las imágenes cambian espontáneamente, sin que nosotros tengamos poder de decisión en el proceso. Únicamente una de las dos imágenes logra acceder a la conciencia, la otra es bloqueada hasta que, por algún evento azaroso, algún disparo solitario de una neurona, se inicia una cascada de eventos que la desbloquea y, por lo tanto, somos conscientes de la otra figura. Pero no vemos “φ”, o al menos eso es lo que dice el consenso científico al respecto.Llegamos a un punto de inflexión interesante, por el momento, apropiado únicamente para un pie de página, pero que luego tomará cada vez más prominencia. Yo escribí “Pero no vemos ‘φ’, o al menos eso es lo que dice el consenso científico al respecto”. Pero ¿qué autoridad tiene el consenso científico respecto de los contenidos de nuestra propia conciencia? Supongamos que hacemos el experimento y vemos constantemente “φ”, y lo comunicamos a nuestros experimentadores. Estamos en una situación complicada: por un lado, no deberíamos estar viendo “φ” (al menos no de acuerdo a la inmensa mayoría de la evidencia experimental existente). Por el otro lado, ¿no somos la autoridad suprema respecto del contenido de nuestra conciencia? ¿Puede el experimentador afirmar que nosotros estamos engañándolo respecto de los contenidos de nuestra propia conciencia? ¿O puede afirmar que nos engañamos a nosotros mismos?
Entonces, esto es lo que (quizás) esperaríamos ver:
FIG. 3.4.3 RIVALIDAD BINOCULAR: EXPECTATIVAS
Y esto es lo que realmente vemos:
FIG. 3.4.4 RIVALIDAD BINOCULAR: REALIDAD
Es posible encontrar neuronas cuya respuesta sea selectiva a cada uno de los dos estímulos. Hay varias formas de hacer esto, pero la más directa es insertar electrodos en distintas zonas de la corteza cerebral de un mono y mostrarle los distintos estímulos por separado (ayuda tener una idea básica de la fisiología del procesamiento visual para saber dónde empezar a buscar). En este caso, el procesamiento visual relacionado con el reconocimiento de objetos y patrones ocurre en regiones que están ubicadas en el lóbulo occipital (región número 3), la corteza extraestriada, y la parte inferior del lóbulo temporal (región número 4).
Una vez que entendemos cómo las distintas células responden a cada estímulo, presentamos una imagen distinta a cada ojo del mono. ¿Tenemos derecho a esperar que la conciencia del mono funcione como la nuestra y que, por lo tanto, el animal vea una sucesión alternante de elipses y palitos en vez de ver “φ” constantemente? Ciertas especies de monos son muy similares a los humanos, y podemos aprovechar esta similitud para inferir el contenido de su conciencia a partir de su comportamiento. Este es el gran legado de los comportamentalistas: métodos para vincular sistemáticamente estímulos con los comportamientos de un animal. Podemos convencer al mono de que apriete un botón u otro, dependiendo del estímulo que está viendo, usando distintas formas de refuerzo positivo (por ejemplo, darle jugo de naranja cada vez que lo hace correctamente). Cuando hacemos eso, aprendemos que la percepción visual de los monos también presenta rivalidad binocular, y entonces podemos proceder con un experimento para buscar los correlatos neuronales de la conciencia visual.
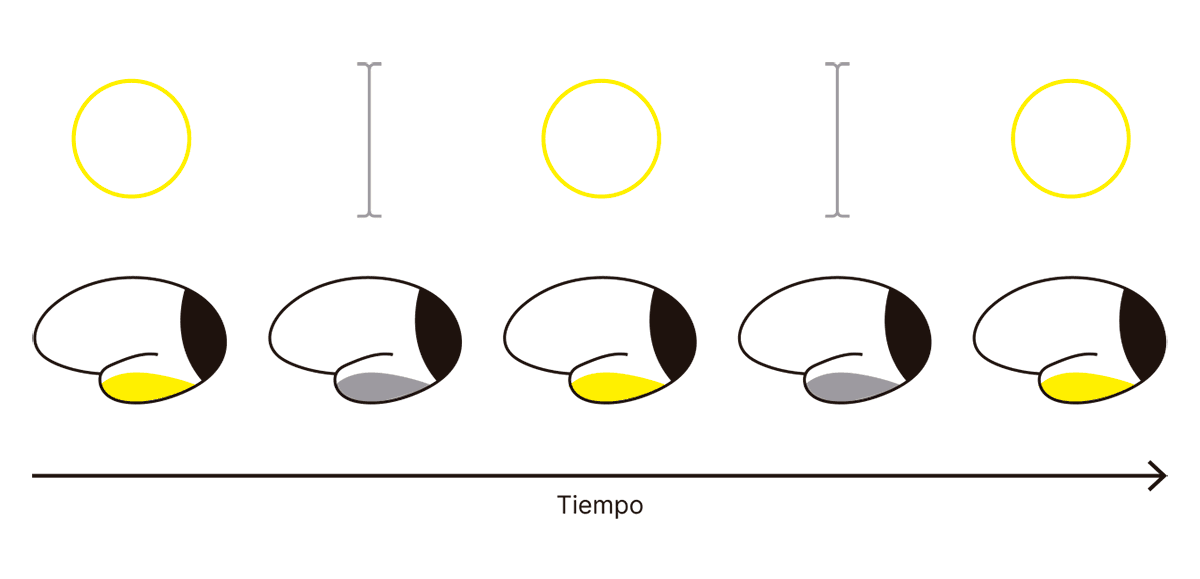
La lógica del experimento es la siguiente: si una neurona mantiene constante su patrón de disparo a lo largo del experimento, entonces no sospechamos que forme parte de los correlatos neuronales de la conciencia visual. Por otra parte, si encontramos neuronas que alternan su patrón de disparo entre aquel correspondiente a la elipse y aquel que corresponde al palito vertical, y si esa alternancia además ocurre con la misma frecuencia con la que se alternan la elipse y el palito vertical en la experiencia consciente, entonces concluimos que esas neuronas están reflejando el contenido de la experiencia consciente del mono:
Este experimento fue realizado por el equipo de Nikos Logothetis en el Instituto Max Planck de Cibernética Biologica, y se encontró que las neuronas correspondientes a las primeras etapas del procesamiento visual en el lóbulo occipital (en color negro en el dibujo) eran indiferentes a la alternancia de los estímulos en la conciencia, mientras que algunas neuronas en la corteza extraestriada y en el lóbulo temporal inferior (que alternan entre amarillo y gris en el dibujoO entre gris claro y gris oscuro para dispositivos blanco y negro.) cambiaban su actividad dependiendo de la imagen presente en la conciencia del mono. Este resultado es extremadamente importante, ya que el lóbulo occipital contiene la corteza visual primaria (abreviada como V1), que es la primera zona de la corteza donde se representa la información visual que proviene de la retina a través del tálamo. De acuerdo al experimento de Logothetis, la corteza visual primaria, entonces, no responde al contenido de la conciencia y, por lo tanto, no puede encontrarse dentro de los correlatos neuronales de la conciencia. Eso no quiere decir que no sea usualmente necesaria para nuestra percepción visual consciente: es tan necesaria para recibir inputs visuales como lo son nuestros ojos y nuestras retinas (o como el teclado, en el caso de una computadora), aunque no esté en el conjunto mínimo de regiones necesarias.
En la corteza visual primaria se realiza el procesamiento elemental de la información presente en la imagen. Primero se extraen los bordes, luego los bordes se combinan en las esquinas, y progresivamente la representación geométrica elemental se vuelve más compleja hasta representar con fidelidad los objetos de la escena visual. Una jerarquía de regiones en el lóbulo occipital y temporal representa otras características relevantes de la imagen, tales como su color o movimiento. Finalmente, la información procede a través de la parte inferior de la corteza temporal, donde se encuentran neuronas cuya activación indica la presencia de objetos específicos, tales como perros labradores, elipses o palitos. Las neuronas de la corteza temporal inferior pueden responder a conceptos en vez de imágenes, es decir, activarse luego de leer las palabras “perro labrador” o de escuchar cierto tipo de ladridos.
El experimento de Logothetis muestra que hay neuronas en las cortezas extraestriada y temporal inferior del mono cuya actividad refleja el contenido consciente, porque sus disparos indican, de forma alternada, la presencia de una elipse y un palito. ¿Significa este resultado que los correlatos neuronales de la conciencia visual están en la actividad de neuronas en estas regiones de la corteza cerebral? Imaginemos que extraemos quirúrgicamente estas regiones del cerebro del mono y las hacemos flotar en un recipiente que les suministra todos los nutrientes que requieren. Además, enviamos señales eléctricas que imitan perfectamente el input que la corteza sensorial recibe de neuronas ubicadas en regiones anteriores del lóbulo occipital. ¿Estamos dispuestos a creer que ese pedazo de tejido cerebral flotando en un tanque es capaz de tener experiencias conscientes por sí mismo? La pregunta es análoga al ejemplo de la cátedra marciana, y ambos pueden resumirse en la misma pregunta: si los correlatos neuronales de una experiencia consciente existiesen y pudiésemos aislarlos, ¿seríamos capaces de creer que por sí mismos bastan para que ocurra una experiencia consciente? Si la respuesta es negativa, entonces quizás sea el momento de aceptar que tal vez haya algo mal con el concepto de correlato neuronal de la conciencia.
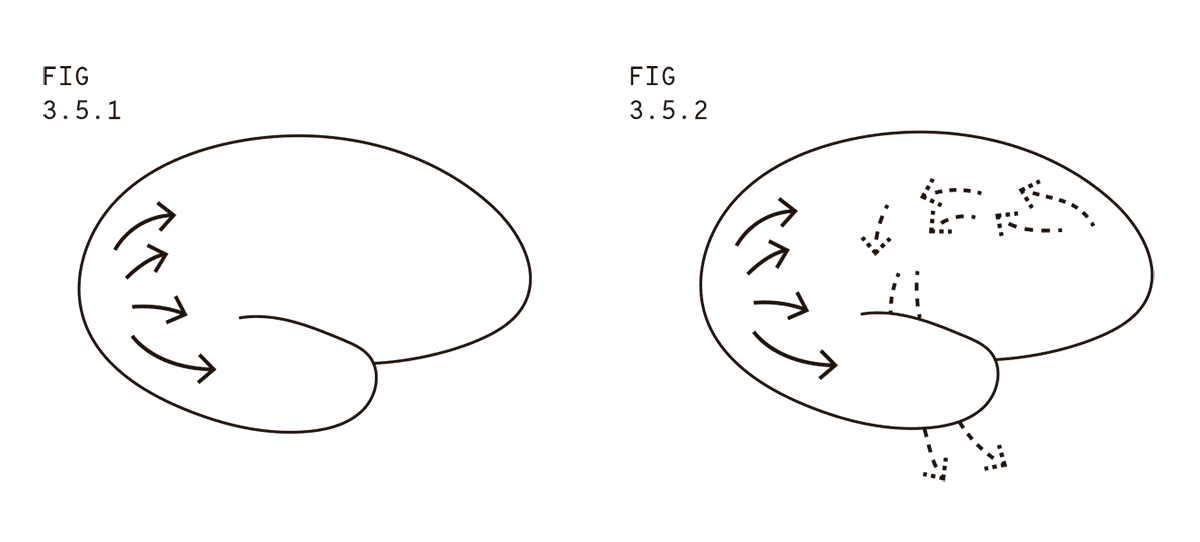
¿Qué tienen en común el experimento marciano y el de Logothetis? Los dos son víctimas de un concepto que heredamos de la tradición comportamentalista: pensar el cerebro como una caja que recibe inputs (por ejemplo, imágenes registradas por la percepción visual) y produce outputs (por ejemplo, reportes hablados o acciones motoras). Si adherimos a esta idea, entonces podemos imaginar que la información sensorial fluye a través del cerebro dirigiéndose a regiones donde es procesada de forma más elaborada, como ilustran las flechas que salen del lóbulo occipital (Fig. 3.5.1).
La información, fluyendo desde la corteza visual primaria, se pregunta: “¿Ya llegamos a la conciencia? ¿Ya llegamos a la conciencia?”. Pero la respuesta es negativa. La información fluye en dirección hacia donde se encuentran los hipotéticos correlatos neuronales de la conciencia, pero todavía no está ahí. Mientras no esté dentro de los correlatos neuronales de la conciencia, entonces, podemos remover esas neuronas y reemplazarlas por una estimulación directa de las neuronas que siguen en la cadena (similar a reactivar una cadena de dominó empujando la pieza siguiente con el dedo en caso de que haya alguna pieza faltante). Si un funcionalista objeta que el sistema artificial con el que determinamos la estimulación debe ser considerado parte del cerebro, contestamos que aún nos encontramos en la periferia cerebral, y que, por lo tanto, podemos determinar la estimulación de muchas otras formas aparte de simulando exactamente el tejido reemplazado.
Por otra parte, el cerebro codifica las acciones motoras mediante neuronas en la corteza motora primaria, una franja de tejido cerebral que está entre el lóbulo frontal (número 1) y el lóbulo parietal (número 2). En primer lugar, se activa una región llamada corteza premotora que, a su vez, activa neuronas ubicadas en la cúspide de la corteza cerebral (conocida como corteza motora suplementaria), las cuales excitan neuronas en la corteza motora. Por último, las neuronas en la corteza motora se proyectan fuera del cerebro a través de la médula espinal para mover nuestros músculos y hacer cosas tales como apretar un botón que diga “perro labrador”. ¿Y de dónde proviene la señal para activar la corteza premotora? Si insistimos en perseguir las flechas que envían la señal de apretar el botón, vamos a encontrar que el antecedente más próximo se encuentra en algún lugar del lóbulo frontal del cerebro (Fig. 3.5.2).
Entradas y salidas del cerebro
Si estamos hablando de un individuo que aprieta un botón que dice “perro labrador” al ver la imagen de un tal perro, entendemos que las flechas de línea continua representan cosas que sucedieron antes que las flechas de líneas discontinuas, porque la información visual tiene que ingresar al cerebro antes de que se produzca una respuesta en reacción a esa información. Además, así como dijimos que las flechas continuas representan el flujo de información que todavía no ingresó a los contenidos de nuestra conciencia, podemos decir que las discontinuas representan información que ya pasó por los contenidos de nuestra conciencia. El motivo es simple: si tomamos la decisión de apretar el botón en base a haber experimentado la imagen de un perro labrador, entonces la actividad motora se genera como causa de algo que ocurrió en nuestra conciencia. Y, por lo tanto, viene después en el tiempo.
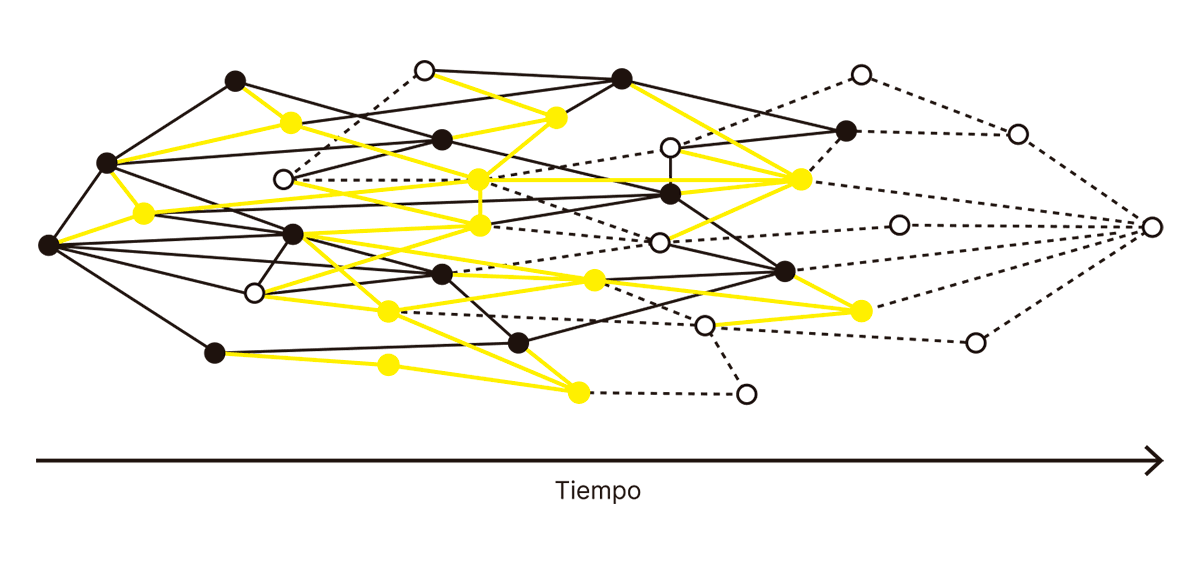
Llegado este punto, es conveniente reemplazar los diagramas de cerebros por otros donde se aprecie con mayor claridad la dimensión temporal de los eventos que están ocurriendo:
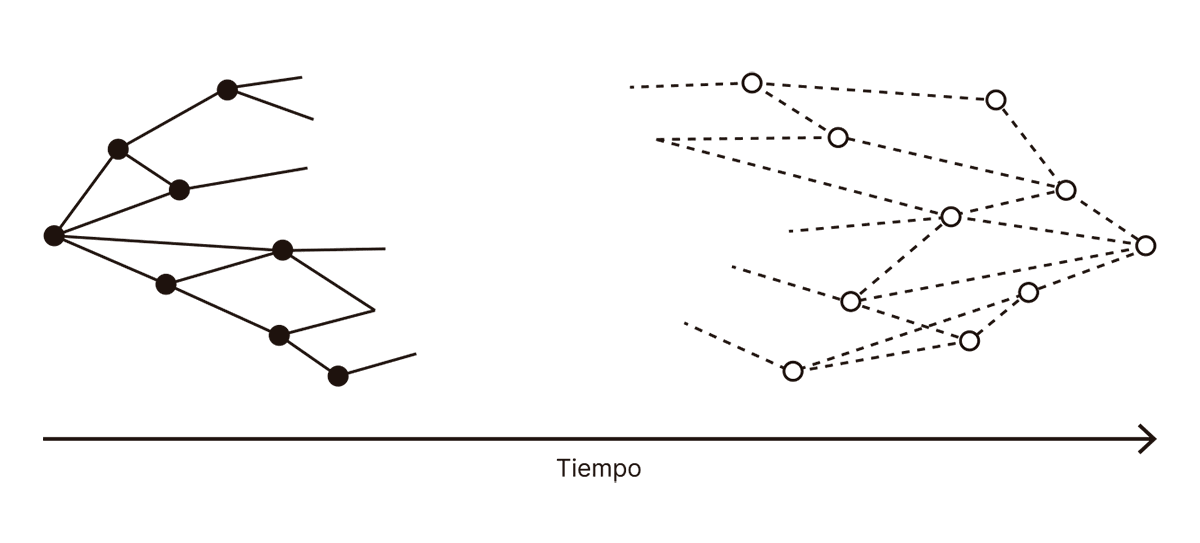
FIG. 3.6.1 ¿CORRELATOS NEURONALES DE LA CONCIENCIA?
El eje horizontal es el tiempo. El comienzo de la flecha corresponde al tiempo de activación de la primera neurona en la corteza visual como respuesta a la imagen del perro labrador. La punta de la flecha se corresponde con el tiempo en el cual se activa la neurona motora que envía la señal al dedo para apretar un botón. Los círculos representan neuronas, y sus colores indican el flujo de información: los círculos y líneas de color negro corresponden a actividad visual que se procesa gradualmente hasta llegar a otras regiones del cerebro que ya no tienen que ver con el procesamiento visual. En cambio, los círculos blancos y las líneas discontinuas representan señales motoras que viajan, por medio de la corteza premotora, la corteza motora suplementaria, y la corteza motora, hacia los músculos en nuestro dedo. En resumen: podemos decir que el color negro corresponde a la trayectoria que realiza el input, y el color blanco, a la trayectoria que realiza el output.
Esto no es más que un esquema, pero es un esquema muy útil: nos dice que concebimos el input visual propagándose hacia adelante en el tiempo, como una secuencia de neuronas en sucesión, similares a una cadena de piezas de dominó que se bifurca y luego converge otra vez, en dirección al espacio vacío en el centro del diagrama. También nos dice que concebimos el output como un proceso similar, pero partiendo del espacio vacío en el centro del diagrama. La pregunta es, entonces: ¿qué hay exactamente en ese espacio vacío?
Ninguna de las neuronas que dibujamos hasta ahora en el diagrama puede formar parte de los correlatos neuronales de la conciencia. Las neuronas negras únicamente propagan una señal hacia donde ocurre realmente la magia de la conciencia; en ese sentido, tienen un rol similar al ojo, a la retina o al nervio óptico. Si estamos convencidos de que la conciencia no está en la retina, también estamos convencidos de que no está en las neuronas negras del diagrama. Lo mismo aplica a las neuronas blancas. Los impulsos nerviosos que activan nuestro dedo son igual de optativos para la conciencia que el movimiento del dedo mismo. Por lo tanto, si estamos convencidos de que la conciencia no está en uno de nuestros dedos,El psicoanalista Jacques Lacan afirmó que el dolor que sentimos luego de golpearnos un dedo del pie contra un mueble no está en el cerebro, sino en el dedo mismo. Pero la pregunta importante no es dónde está el dolor, sino dónde está el dedo: así como tenemos un dedo físico que se mueve y se lastima, ese mismo dedo está representado múltiples veces dentro de nuestro cerebro, y el dolor está en alguno de esos dedos. Parafraseando a Paul Éluard (“Hay otros mundos, pero están en este”), diríamos que hay otros dedos, pero están en el cerebro. también estamos convencidos de que no está en las neuronas blancas del diagrama.
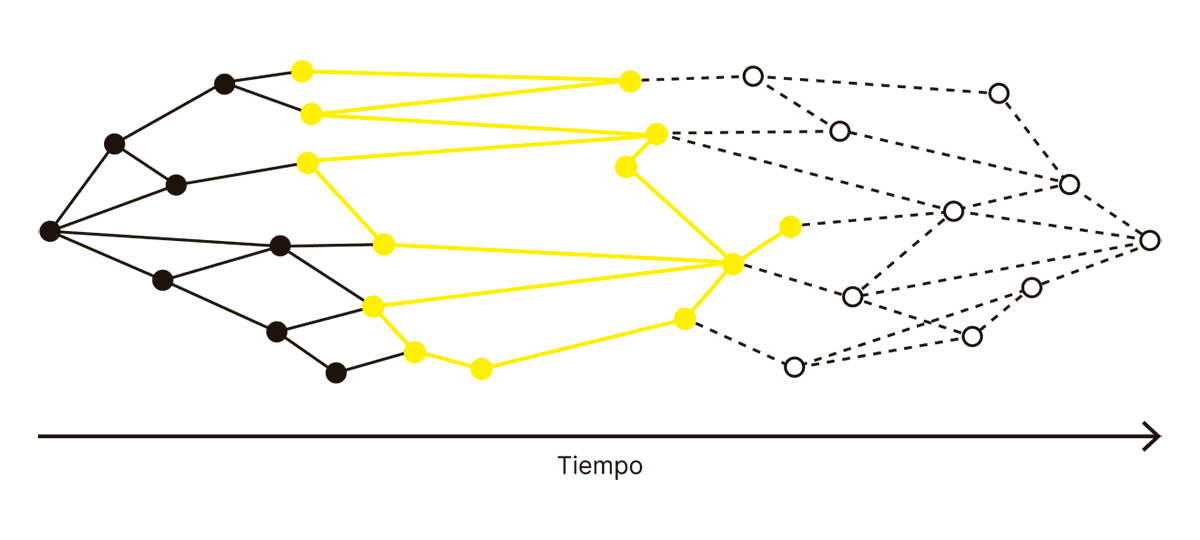
Pero en algún lugar del espacio vacío, el negro debe transformarse en blanco. Esa es la cúspide del procesamiento cerebral, el momento en el cual el input se transforma en output, y la sensación se transforma en acción. Crick y Koch imaginaron cómo las flechas negras convergen en un grupo de neuronas que no son ni negras ni blancas, neuronas donde realmente ocurre la magia, donde encontramos los correlatos neuronales de la conciencia:
FIG. 3.6.2 ¿CORRELATOS NEURONALES DE LA CONCIENCIA?
Si las neuronas amarillas son realmente el correlato neuronal de nuestra experiencia consciente, entonces (por definición) son el mínimo conjunto de neuronas cuya activación es necesaria para tener esa experiencia. Podríamos tomar un bisturí y cortar el resto del cerebro salvo la porción que las contiene, y si suministramos los inputs correctos de forma externa, ese conjunto de neuronas aisladas tendría que poseer una experiencia consciente:
FIG. 3.6.3 ¿CORRELATOS NEURONALES DE LA CONCIENCIA?
Llegamos nuevamente a un lugar familiar. Encontramos un conjunto de neuronas que constituyen lo mínimo indispensable para que se manifieste la conciencia. Pero esas neuronas no pueden hablar o moverse (porque la conciencia no necesita de habla ni de movimiento). Esas neuronas no tienen memoria, ni emociones, ni lenguaje; no pueden comunicarse con el mundo y decir “es un perro labrador”, tampoco pueden mover un dedo o pensar en mover un dedo, porque todas estas cosas son, al menos en apariencia, innecesarias para la conciencia. Lo único que esas neuronas pueden hacer es (en caso de recibir los inputs adecuados) experimentar subjetivamente la imagen de un perro labrador. Esas neuronas y su actividad son, en un sentido prácticamente literal, la manifestación física más pura de la experiencia consciente de percibir un perro labrador.
Podemos creer en esta imagen que Crick y Koch conjuran para nosotros. Podemos creer en científicos marcianos que reducen el cerebro a un racimo de neuronas cuya actividad constituye conciencia pura flotando en el vacío; podemos creer que seccionar la corteza temporal inferior de un mono, meterla en un frasco con nutrientes y alimentarla con impulsos eléctricos preserva la conciencia visual del mono; podemos estar convencidos de todas estas cosas, aunque desafíen en gran medida nuestro sentido común. Lo que seguro no podemos es verificar que estas neuronas tienen la experiencia consciente que podríamos afirmar que poseen. Una vez eliminadas las innecesarias neuronas que producen un reporte (hablar, apretar un botón), los correlatos neuronales de la conciencia son como el interior de un agujero negro: completamente incognoscibles e inaccesibles para los observadores externos, aislados causalmente del resto de las cosas y seres de este universo, privados en el sentido más intenso de la palabra. No podemos verificar que el racimo de neuronas amarillas posea conciencia y, por lo tanto, parecemos habernos topado con una frontera de la ciencia.
Aunque quizás sospechen, como yo sospecho, que hay algo extraño flotando alrededor de estos diagramas que estoy dibujando. Estoy asumiendo una cierta simplicidad que jamás vamos a encontrar en un objeto tan complejo como el cerebro. El hecho de que la actividad ascienda en el tiempo propagándose en neuronas negras y termine el proceso propagándose en neuronas blancas no implica automáticamente que existe un lugar bien definido donde ocurre la mágica permutación de los colores que llamamos correlato neuronal de la conciencia.
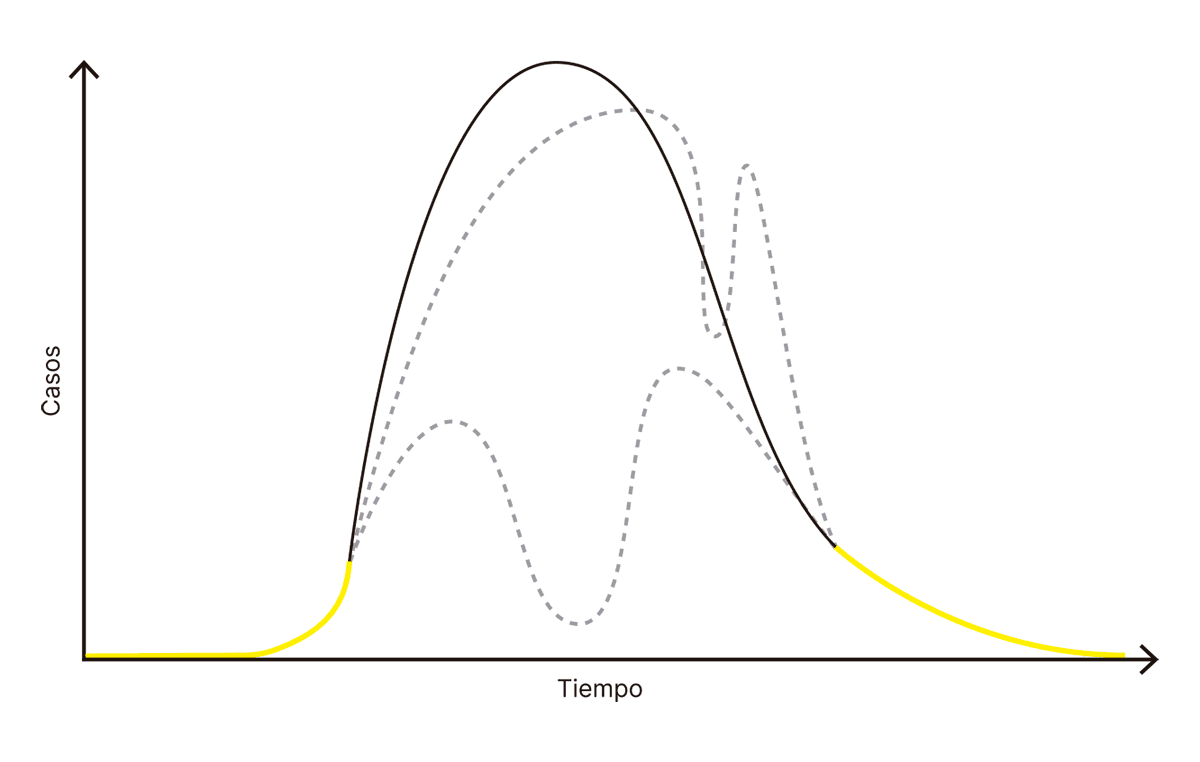
Estoy escribiendo estas páginas en plena evolución de la pandemia COVID-19, un evento terrible que nos mantiene confinados en cuarentena, mientras miramos impotentes cómo sube el número de casos. Hoy, 28 de julio de 2020, se registraron 5939 casos nuevos en Argentina y (descartando algunas fluctuaciones irrelevantes) esperamos que el número siga subiendo. Pero al mismo tiempo sabemos que la pandemia COVID-19 va a terminar algún día. Ese día, vamos a empezar a ver cómo la cantidad de nuevos contagios desciende gradualmente hasta llegar a cero. Empezamos, entonces, con una función creciente de casos y terminaremos con una función decreciente de casos, por lo tanto asumimos que en el medio vamos a encontrar un pico de casos: un día en el cual la cantidad de contagios será máxima, superior tanto al día anterior como al siguiente.
Pero en realidad no tiene por qué ser así.Me encuentro editando la primera versión del manuscrito durante el día 24 de enero de 2021 y puedo, efectivamente, confirmar que el escenario con un solo pico bien definido era una ingenuidad. Cualquier curva que tracemos que comience y termine como los dos trazos amarillos representa una evolución posible (aunque no necesariamente probable) de la pandemia:
FIG. 3.6.4 FUNCIÓN DE ESTADO DE LA PANDEMIA
La complejidad de algunas de las curvas que dibujé es quizás excesiva. Pero también podríamos argumentar que la simplicidad de la curva negra (con un pico único) también es excesiva. Podemos equivocarnos profundamente si esperamos siempre encontrarnos con el escenario más sencillo compatible con lo que sabemos sobre el comienzo y el final de un proceso.
Lo mismo sucede en el caso del cerebro. Si bien las cosas son ordenadas tanto en el comienzo como en el final, cuando las neuronas negras entran en contacto con las neuronas amarillas se entrelazan en un nudo de complejidad inimaginable. Pensamos en remover las neuronas negras que están en el borde del nudo, pero cada neurona que quitamos reconfigura el flujo de información y cambia el color de todas las neuronas que se encuentran alrededor y dentro del nudo. Simultáneamente, encontramos neuronas que transmiten inputs, outputs y neuronas que no hacen ninguna de las dos cosas. Cuando el científico marciano destruyó casi todas las neuronas y anunció haber encontrado los “correlatos neuronales de la conciencia”, su clase estalló en carcajadas porque todos entendieron que, como consecuencia del proceso mismo, las neuronas restantes ya no tenían nada que ver con la conciencia.
El cerebro es un sistema inmensamente complejo de circuitos neuronales entrelazados. Algunos de ellos cumplen funciones más o menos identificables; por ejemplo, representar información sensorial de distintos tipos, amplificar cierto tipo de información, acumular evidencia para la toma de decisiones, consolidar distintos tipos de memoria, etc. Lo que seguro no vamos a encontrar en el cerebro es un grupo de neuronas aislado donde ocurra la magia de la conciencia, un circuito donde se represente la información sensorial tal como aparece en nuestra experiencia subjetiva. Esto sería esperable si la conciencia estuviese en la cúspide de la jerarquía del cerebro, en un sitio desde el cual emana nuestra voluntad, tal como Descartes imaginó la glándula pineal. En realidad, la conciencia siempre involucra distintas funciones del cerebro (aquello de lo que hablamos o escribimos, lo que recordamos, lo que sentimos) y es inseparable de estas funciones. Intentar aislar los correlatos neuronales de la conciencia es como intentar separar los polos de un imán: aun si rompemos el imán en dos mitades, cada mitad pasará a tener dos polos en el mismo pedazo.
El escenario que enfrentamos se corresponde con el siguiente diagrama:
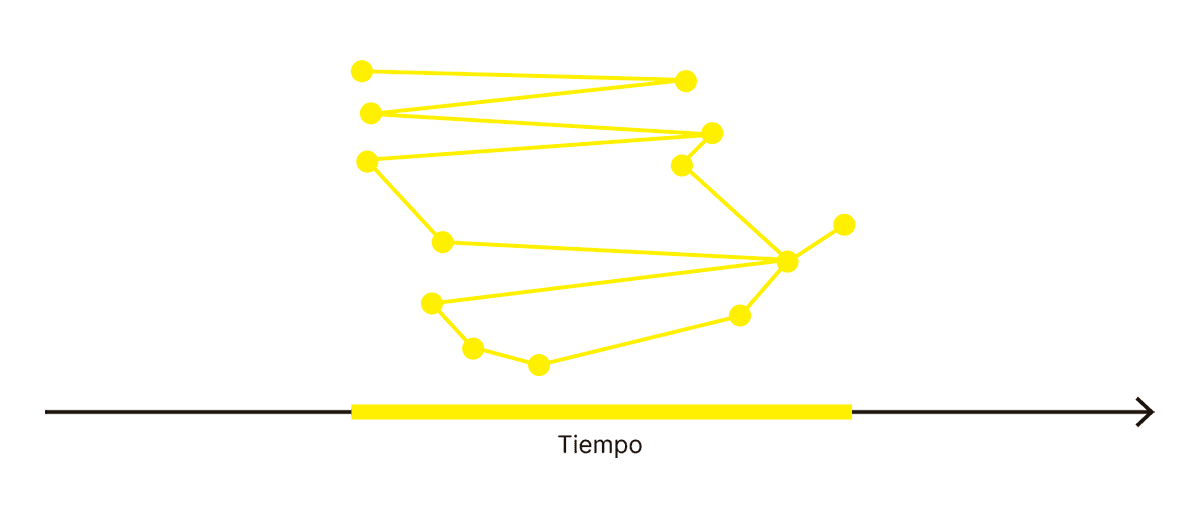
FIG. 3.6.5 ¿CORRELATOS NEURONALES DE LA CONCIENCIA?
En el centro, las neuronas que no pertenecen claramente a la entrada ni a la salida del sistema se encuentran distribuidas en múltiples funciones, sin una frontera clara entre el flujo hacia el interior y el exterior del cerebro. No hay una jerarquía a la cual ascender para encontrar en su cumbre la conciencia; en vez de eso, la conciencia surge de procesos neuronales distribuidos y entrelazados íntimamente con el tejido de las funciones mentales. Por lo tanto, es imposible hacer cortes para separar la conciencia sin deshacer el entramado.
Según cuenta la leyenda, un oráculo profetizó que el próximo hombre en entrar a la ciudad de Frigia (hoy una región de Turquía occidental) en un carro tirado por bueyes sería el próximo rey. Un campesino llamado Gordias entró a la ciudad en un carro, tal como había predicho el oráculo, y así fue coronado rey de Frigia. Luego de los festejos y presa de una enorme gratitud, su hijo Midas ofrendó el carro al dios Zeus atándolo a un poste con un nudo tan intrincado que sus cabos se escondían en su interior, al punto que era imposible identificar el trayecto de la soga. Se dijo, entonces, que aquel que fuera capaz de desatar el nudo hecho por el hijo de Gordias sería coronado emperador de toda Asia.
Luego de muchos años, en el siglo IV a. C., Alejandro Magno se dirigía al Oriente hacia la guerra con el Imperio persa, cuando su camino lo acercó a la ciudad donde el nudo gordiano seguía atado. Fue entonces cuando se apersonó para hacer pública su intención de conquistar todo el continente. Ante la mirada expectante de la ciudad, Alejandro Magno se acercó al nudo e intentó desatarlo, pero sus dedos se deslizaban sin encontrar la forma de mover la cuerda. Durante horas, intentó en vano deshacer el nudo de Gordias mientras sus asesores y generales se miraban de reojo, nerviosos. Pero al caer la noche desenvainó su espada y cortó el nudo por la mitad, declarando que “cortarlo es lo mismo que desatarlo”. Entonces todos los presentes se postraron ante la presencia del futuro emperador de Asia, Alejandro Magno, el hombre que había conseguido desatar el nudo gordiano.
La idea de que existe tal cosa como un correlato neuronal de la conciencia es una burda simplificación, el equivalente neurocientífico de cortar el nudo gordiano de la conciencia. Nos presenta una imagen del cerebro donde las neuronas indispensables para la conciencia pueden separarse prolijamente de aquellas que son optativas, y sugiere que existen límites bien definidos en los procesos que ocurren antes y después de que comience el proceso de la percepción consciente. También implica que podemos desatar el nudo simplemente cortando el cerebro a través de esos límites, logrando aislar una porción de materia que corresponde a un puro fragmento de contenido consciente, y nada más.
A pesar de su simplicidad conceptual, le debemos a esta idea el nacimiento de la neurociencia contemporánea de la conciencia. Quizás porque la idea es engañosamente fácil de entender, disparó una enorme cantidad de investigaciones sobre la relación entre el cerebro y la conciencia. Y quizás si algunos de los problemas que discutimos en este capítulo hubiesen estado claros desde el principio, la historia habría sido diferente.
Arthur Schopenhauer escribió sobre el problema de la subjetividad, la identidad y la voluntad, y lo denominó el “nudo del mundo” (Weltknoten). Cuando intentamos imaginar la complejidad con la que se mezclan los inputs y los outputs en el cerebro, esta metáfora parece ser adecuada. No existe mucha controversia sobre las neuronas que corresponden a la entrada del sistema y aquellas que corresponden a la salida. No están realmente “anudadas” y, por lo tanto, no están implicadas en la conciencia, al menos no más que la pantalla de nuestro monitor, nuestros lentes o nuestras retinas; no más que el volante del auto, los guantes que llevamos puestos o los músculos de nuestras manos. Pero a medida que avanzamos y retrocedemos hacia el gran nudo en el centro de la mente, las cosas dejan de ser tan claras.
Como veremos en el próximo capítulo, el borde del nudo es el campo de batalla donde se enfrentan distintas teorías de la conciencia. Una de las principales batallas consiste en determinar qué se encuentra dentro y qué se encuentra fuera del nudo, es decir, dónde comienza y dónde termina el antes y el después de la conciencia. No hay respuestas fáciles, porque el territorio no es una línea recta, sino una frontera de complejidad inimaginable. Enfocados en cartografiar los bordes de este nudo, los neurocientíficos de la conciencia tienden a evitar una pregunta sencilla pero fundamental, una pregunta que tiene que cambiar por completo la forma en que abordamos el problema de la conciencia: ¿cuál es la probabilidad de que dos personas distintas posean un mismo nudo, con los mismos bordes, exactamente idénticos en su complejidad? En un lenguaje menos poético, ¿podemos asumir una frontera universal que separe los procesos pre- y postconscientes, válida para todos los seres humanos del planeta, o al menos para la mayoría de ellos? Los neurocientíficos contemporáneos se comportan como si pudiesen (y discuten acaloradamente al respecto cuando no lo logran). Pero dadas las idiosincrasias de cada cerebro humano particular, es razonable arriesgar una respuesta negativa. Y en ese caso, ¿cuáles son las implicaciones de esa individualidad para el entendimiento científico de la conciencia?
¿Qué pasó finalmente con la propuesta de Crick y Koch? Muchos neurocientíficos que hoy investigamos la conciencia desconfiamos profundamente de la posibilidad de encontrar correlatos neuronales de experiencias conscientes. Eso no significa que no apliquemos toda la tecnología a nuestra disposición para investigar la relación que existe entre el cerebro y la experiencia subjetiva. Experimentos como el de Logothetis poseen un gran valor: si bien los correlatos neuronales de la conciencia no se encuentran en la corteza temporal inferior, al menos sabemos que la actividad en esa región está implicada de alguna forma con los contenidos de la conciencia, mientras que otras regiones (tales como la corteza visual primaria) parecen ser optativas.
Luego de la propuesta original de Crick y Koch, una larga lista de candidatos para los correlatos neuronales de la conciencia desfilaron por las revistas más prestigiosas de neurociencia: oscilaciones corticales y tálamo-corticales a 40 Hz, bucles de conexiones tálamo-corticales, actividad en los núcleos talámicos intralaminares, actividad de neuronas en la corteza temporal inferior, formaciones de colaciones neuronales en la corteza frontal y muchas otras. Todas las propuestas fueron descartadas por la evidencia experimental, es decir, en todos los casos se demostró que o bien era posible encontrar conciencia en ausencia del mecanismo candidato, o bien el mecanismo candidato era insuficiente por sí mismo para generar experiencias conscientes.
Incluso se ha cuestionado que la corteza visual primaria no se encuentre involucrada en los correlatos neuronales de la conciencia. Cierto tipo de lesiones en esa zona del cerebro dan lugar a un fenómeno muy curioso llamado visión ciega. Los sujetos con visión ciega son incapaces de reportar los contenidos de su experiencia visual. Si le preguntamos a alguno de ellos sobre la imagen de un perro labrador, negará enfáticamente estar viendo un perro. Pero si insistimos hasta que el sujeto se anime a adivinar qué es lo que le estamos mostrando, vamos a obtener la respuesta “perro” con una mayor frecuencia de la esperada por puro azar. Una interpretación posible es que dañar la corteza visual primaria no limita el flujo de información hacia otras regiones del cerebro (al fin y al cabo, la información tiene que llegar a zonas relacionadas con la producción del lenguaje para que el sujeto pueda adivinar con un nivel superior al azar), pero sí limita directamente los contenidos de la conciencia, y por lo tanto se encuentra involucrada con los correlatos neuronales de esta. Existen múltiples argumentos en contra de esta observación, pero uno de ellos en particular (que debemos a David Chalmers) es consistente con nuestra preocupación por esa estructura que llamamos el nudo de la conciencia. Chalmers considera la posibilidad de que la corteza visual primaria esté lo suficientemente “anudada” como para que dañarla modifique y reorganice completamente los correlatos neuronales de la conciencia visual.
Finalmente, el estudio de la conciencia siguió por el carril que Crick y Koch habían descartado en su propuesta original: el de las ciencias cognitivas y sus explicaciones en términos funcionales. En su ambición por encontrar el equivalente físico de una experiencia consciente pura, desestimaron el papel que los reportes explícitos juegan a la hora de determinar la existencia de esa experiencia consciente. Los monos se pueden entrenar para realizar tareas simples y pasivas, como la rivalidad binocular y otros experimentos similares; exigirles una participación activa en los experimentos (por ejemplo, un reporte explícito) es considerablemente más difícil. Tenemos suficiente familiaridad con ciertas especies de mono como para interpretar sus comportamientos como la clase de comportamientos que nosotros mismos haríamos en respuesta a ciertas experiencias conscientes.Esto explica por qué no vemos experimentos sobre las bases neuronales de la conciencia hechos en ratas, gatos o perros. No dudamos de que estos animales posean un mundo interior, con experiencias subjetivas de distinto tipo e intensidad. El problema es que no podemos establecer un código para comunicar unívocamente los contenidos de esa conciencia. Para ventaja nuestra (y desventaja suya), es mucho más fácil relacionarnos e identificarnos con el comportamiento de un mono. Pero cuando revisamos los trabajos con primates no humanos, encontramos un problema común: la incapacidad de obtener un reporte verbal claro y preciso impide delinear con claridad el límite de los procesos que ocurren antes y después de la conciencia.
Una forma de resolver este problema es investigando las bases neuronales de la conciencia en seres humanos. Si bien no podemos insertar electrodos a voluntad en el cerebro de personas sanas, existen técnicas de neuroimágenes capaces de informarnos con detalle sobre los cambios eléctricos y metabólicos que ocurren en paralelo con una experiencia reportada como consciente, en comparación con una experiencia reportada como no consciente. Y si bien es cierto que el lenguaje parece ser un proceso optativo en cuanto a la experiencia consciente, el reporte explícito nos permite estimar con mayor precisión los límites del nudo, aun si los sobrestimamos sistemáticamente en la dirección posterior a la conciencia. No hay ambigüedad posible ante una persona que nos mira a los ojos y nos dice “Es un perro labrador”.
En resumen, la idea de Crick y Koch de encontrar mecanismos neuronales asociados con la conciencia (independientemente de su rol funcional en el cerebro) estaba equivocada. Además, desestimaron que el estudio de la conciencia en humanos puede verse enormemente facilitado por las distintas funciones cognitivas que les permiten relacionarse con otros humanos (los experimentadores) y comunicarse sin ambigüedades. El reporte explícito es una herramienta útil para delimitar los delicados contornos del nudo de la conciencia, estableciendo fronteras en las que podemos confiar tanto como confiamos en los reportes de nuestros sujetos experimentales.
Al final, no estaríamos tan equivocados en incorporar al diagrama un cuadradito que diga “conciencia”:
FIG. 3.7 DE LA LLUVIA A LA DECISIÓN, PASANDO POR LA CONCIENCIA
Entendemos que la conciencia se puede estudiar en términos funcionales, es decir, abandonando la idea de que es posible aislar “conciencia pura” en un fragmento de cerebro, y por lo tanto abordamos el problema de explicarla en términos de su rol funcional. El desafío es entender qué sucede dentro del cuadradito, es decir, identificar el rol de la conciencia en el procesamiento de información en el cerebro. En el próximo capítulo, la discusión sobre cuál es exactamente este rol nos va a llevar a la teoría más exitosa que tenemos al día de hoy sobre la conciencia.
¿De qué está hecha la experiencia? ¿En qué lugar de nuestro cerebro reside el sabor de una comida o el sonido de nuestra canción favorita? Un tratado que condensa una década de investigación original del Dr. Enzo Tagliazucchi, y discute las concepciones históricas y contemporáneas sobre la conciencia humana, uno de los enigmas más insondables para la humanidad.
464 páginas, con gráficos a color. Ganador del Sello Buen Diseño.
El nudo de la conciencia$37.500
¿De qué está hecha la experiencia? ¿En qué lugar de nuestro cerebro reside el sabor de una comida o el sonido de nuestra canción favorita? Un tratado que condensa una década de investigación original del Dr. Enzo Tagliazucchi, y discute las concepciones históricas y contemporáneas sobre la conciencia humana, uno de los enigmas más insondables para la humanidad.
464 páginas, con gráficos a color. Ganador del Sello Buen Diseño.
El nudo de la conciencia Ebook$15.000
¿De qué está hecha la experiencia? ¿En qué lugar de nuestro cerebro reside el sabor de una comida o el sonido de nuestra canción favorita? Un tratado que condensa una década de investigación original del Dr. Enzo Tagliazucchi, y discute las concepciones históricas y contemporáneas sobre la conciencia humana, uno de los enigmas más insondables para la humanidad.
Formato .epub
El nudo de la conciencia - edición especial$45.000
¿De qué está hecha la experiencia? ¿En qué lugar de nuestro cerebro reside el sabor de una comida o el sonido de nuestra canción favorita? Un tratado que condensa una década de investigación original del Dr. Enzo Tagliazucchi, y discute las concepciones históricas y contemporáneas sobre la conciencia humana, uno de los enigmas más insondables para la humanidad.
464 páginas, con gráficos a color. Edición especial con tapa dura.
Ciencia y diseño para entender el mundo como es, cómo podría ser y cómo queremos que sea.